
- Home
- Matemática Aurea
- Flor de la Vida
- Sólidos Sagrados
- Geometría sagrada y energía
- Y además...
- Modelos 3D
- Flor de la Vida 3D
- Sólidos Platónicos 3D
- Sólidos Arquimedianos 3D
- Sólidos de Catalan 3D
- Dodecaedro Disdiakis 3D
- Dodecaedro Pentakis 3D
- Dodecaedro Rómbico 3D
- Hexaedro Tetrakis 3D
- Hexecontaedro Deltoidal 3D
- Hexecontaedro Pentagonal 3D
- Icosaedro Triakis 3D
- Icositetraedro Pentagonal 3D
- Icositetraedro deltoidal 3D
- Octaedro triakis 3D
- Tetraedro Triakis 3D
- Triacontaedro Disdiakis 3D
- Triacontaedro Rómbico 3D
- Pirámides y Prismas 3D
- Estelaciones 3D
- Phi en ...
- Sólidos Sagrados en ...
- Música y geometría
- Acerca de
- Modelos 3D
Phi en la estructura del ADN
1.- Introducción
Este artículo trata sobre las dimensiones de la molécula del ADN según las investigaciones de Mark E. Curtis [1]. Es ampliamente conocido que cada giro completo de la doble hélice del ADN mide 21 Å de ancho por 34 Å de largo. 34 y 21 son dos números consecutivos en la sucesión de Fibonacci. Por lo tanto su cociente, 1.6190476, aproxima de cerca la Razón Aurea φ=1.6180339...
Hace casi 20 años Mark E. Curtis empezó una investigación de la estructura del ADN con la intención de producir una serie de dibujos y pinturasde la doble hélice [1]. Se embarcó en dibujos a escala de la estructura en hélice usando las dimensiones estándar de los libros de texto que se derivan de los datos de difracción de Rayos X. En el transcurso de su trabajo, surgieron discrepancias, y resultó claro para él quela estructura de Crick y Watson [2] no seguía principios geométricos. Además sus intentos de trasladar su teoría de 2D a 3D chocaron con problemas topológicos considerables. Él descubrió que una minoría creciente reconoce defectos en las conclusiones de Crick y Watson, específicamente en términos de su topología y por lo tanto la habilidad de la estructura para replicarse a sí misma. Aunque algunas investigaciones han ido tan lejos hasta para cuestionar la propia existencia de la estructura en doble hélice, su propuesta retiene la doble hélice como una base fundamental.
Sin comprometer la esencia de su estructura, Mark E. Curtis propuso una solución a las inconsistencias geométricas mediante un simple cambio en la posición de alineamiento entre las purinas y las pirimidinas. Este realineamiento está basado por completo en principios geométricos. Además, un conjunto de relaciones matemáticas sencillas relacionadas con la Geometría Sagrada describen una hélice tridimensional que se adecúa a las medidas conocidas del ADN. Esta relectura del ADN genera una gran pureza y simplicidad cuando se aplica a la estructura molecular de las bases.
2.- Geometría Sagrada bajo la molécula de ADN
Aunque en teoría es posible construir una hélice partiendo de cualquier serie de polígonos, resulta haber sólo una formación poligonal que cumpla totods los criterios requeridos: diez pentágonos rgulares orientados según un decágono (Figura 1).
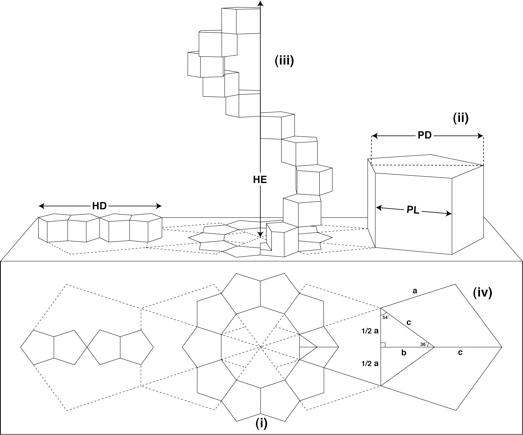 |
| Figura 1: Estructura geométrica del ADN basada en la Geometría sagrada (reproducido de [1]): |
Cuando se traduce a 3D, estos pentágonos se convierten en prismas con todas las longitudes iguales (ii).Una vuelta completa de la hélice está formada por la rotación y extensión progresiva de diez prismas pentagonales regulares (iii). El diámetro de la hélice (HD) y la altura de la extensión de la hélice (HE) tienen una razón constante y proporcional con el lado pentágono (PL) y su altura (PD).
De la figura precedente, PL=a y PD=b+c obedecen a la expresión siguiente:
a partirde la cual los valores exactos de PD, HD y HE se pueden calcular numéricamente:
PD = 1.5388417686·PL
HD = 4·PD = 6.1553670744·PL
HE = 10·PL
Vamos a aplicar una de las dimensiones conocidas del ADN a las ecuaciones anteriores. La anchura de la base se sabe que mide aproximadamente PL=3.4Å. Así pues tenemos
PD = 1.5388417686·PL = 5.23206201324 Å
HD = 6.1553670744·PL = 20.928248053 Å
HE = 10·PL = 34 Å
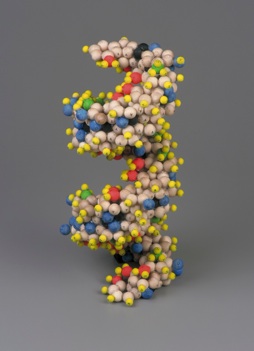
Estas cifras parecen coherentes con las dimensiones conocidas del ADN reveladas por los datos de difracción de rayos X. A partir de ellas, Mark E. Curtis construyó el modelo tridimensional exacto de la doble hélice del ADN mostrado en la (Figure 2). Utilizó simples prismas pentagonales para ilustrar la hélice tridimensional. Para asegurar la estabilidad de la estructura como existiría en realidad, es necesario poner pentágonos adyacentes en cada plano y luego apilarlos consecuentemente, creando así la doble hélice:
 |
 |
| Figura 2: Modelo geométrico de la doble hélice del ADN por Mark E. Curtis (reproducido de [1]) | |
3.- Aplicación a la estructura molecular del ADN
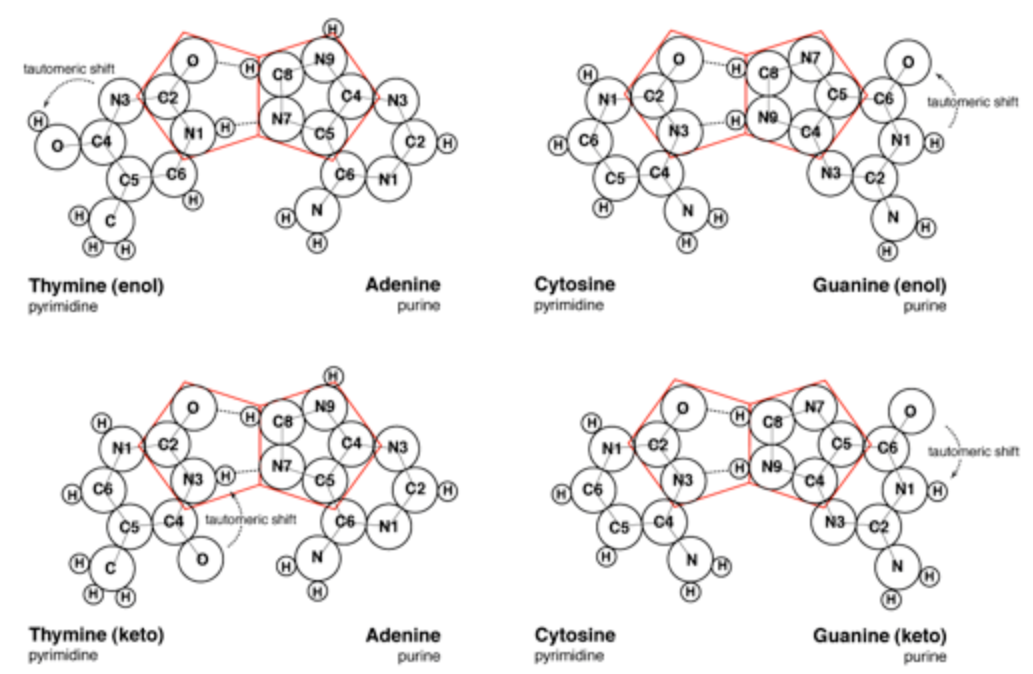
La unión de purinas y pirimidinas en la propuesta de Crick y Watson localiza los pentágonos moleculares que forman parte de las purinas en la parte exterior de las bases apareadas (Figura 3). Sin embargo, se ha demostrado que una geometría más fluida y natural requeriría pentágonos adyacentes en el corazón de la unión de bases. Mark E. Curtis propone reorientar la unión moluecular establecida de forma que los pentágonos de la adenina (A) y guanina (G) fromen enlaces de hidrógeno con las estructuras hexagonales de la timina (T) y la citosina (C) (Figura 4). Esta distribución espacial de los enlaces de hidrógeno hace viable el segundo pentágono adyacente. Además, también encaja con nuestra comprensión de los enlaces moleculares y mantiene la especificidad de G conh C y A con T.
 |
|
|
(a) Unión de bases según Crick y Watson [1]. |
|
 |
|
|
(b) Uniones pentagonales propuestas por M.E. Curtis [1]. |
|
| Figura 3 |
En este punto Mark E. Curtis hace referencia a las diferentes formaciones que pueden adquirir las moleculas de guanina y timina, que son los estados enol y keto. La Guanina y la timine pueden existir en cualquiera de las dos configuraciones y se cree que passan por cambios tautoméricos de la una a la otra. En otras palabras, un salto indiscernible y espontáneo por parte del átomo de hidrógeno desde un átomo específico de oxígeno hacia un átomo específico de nitrógeno, o viceversa. Mientras que la propuesta de Crick y Watson requiere la forma keto, la propuesta alternativa de Curtis es viable en cualquiera de las dos formas.
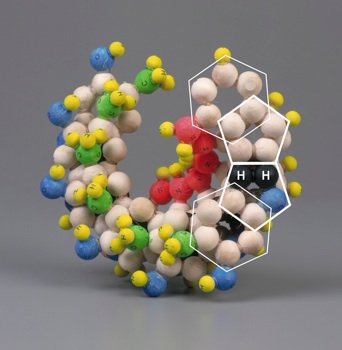 |
 |
| Figura 4: Construcción tridimensional para mostrar las modificaciones de las bases propuesta por Mark E. Curtis [1]. | |
4.- Implicaciones del modelo
La estructura del ADN propuesta por Mark E. Curtis está completamente basada en principios matemáticos. Aunque las modificaciones geométricas a las uniones entre bases es relativamente menor, la doble hélice resultante manifiesta una claridad muy distinta respecto a la propuesta de Crick y Watson, y parece aportar luz en un conjunto de áreas en continua incertidubre.
Las implicaciones de estos hallazgos son inevitablemente de largo alcance y potencialmente podrían afectar muchas areas de investigación. Las características principales de esta propuesta son:
- Ecuaciones geométricas que predicen las dimensiones de la estructura del ADN. La geometría pentagonal no sólopredice las dimensiones de la hélice, sino que demuestra también un "principio causativo".
- La geometría pentagonal proporciona la dinámica necesaria para construir una estructura en forma de hélice consistente, estable y uniforme, y también establece por qué deben haber diez bases contenidas dentro de una sola vuenta de la hélice. Curtis predice pequeños grados de variación cuando se traslade esta estructura a nivel molecular, del orden de 9.5 a 10.5 bases por giro o quizá más.
- Tanto el centro hueco como la formación estructural lado con lado aseguran un acceso instantáneo a cualquier punto dentro de la hélice. Esto permitiría al ADN (incluso circular) abrirse y cerrarse durante sus funciones de replicación sin enredarse consigo mismo.
- Las modificaciones en ls uniones entre bases pueden existir tanto en la forma enol como keto.
- Aunque los esqueletos de azúcar-fosfato sin duda se demostrarán integrales para la estabilidad de la estructura en forma de hélice, la geometría de las propias moléculas de pares de bases es la responsable última de la formación de la hélice.
5.- Referencias
[1] Mark E. Curtis, "The geometry of DNA: a structural revision".
[2] Crick, F. H. C. & Watson, J. D. (1953) Nature, vol 171, 737-738.